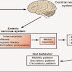









'Εκκριση Γαστρικού Οξέος
Για να ξεκινήσει η έκκριση γαστρικού
οξέος θα πρέπει να ενεργοποιηθεί η αντλία H+ - K+ ATPase.
Η ενεργοποίηση της αντλίας προκαλείται ως επακόλουθο 2 συνεργικών καταστάσεων
την αύξηση ενδοκυττάριας συγκέντρωσης Ca2+
και cAMP
με τελικό αποτέλεσμα την ενεργοποίηση μιας σειράς από πρωτεϊνικές κινάσες οι οποίες
φωσφορυλιώνουν πρωτεΐνες και θέτουν σε λειτουργία την αντλία.(Για περισσότερες πληροφορίες δες Κυτταρικοίμηχανισμοί έκκρισης γαστρικού οξέος
 Σε δεύτερη φάση τα πεπτίδια
είναι ικανά να αλληλοεπηρεάσουν το ένα με
το άλλο ,έτσι ώστε η έκκριση του ενός να
θεωρείται ως διεγέρτης για την έκκριση
κάποιου άλλου. Όπως θα δούμε στη συνέχεια στη κεφαλική φάση των γαστρικών
εκκρίσεων όπου δεν έχει φτάσει ακόμη η τροφή στο στομάχι η έναρξη παραγωγής του
γαστρικού οξέος γίνεται ως αποτέλεσμα διέγερσης του πνευμονογαστρικού.
Σε δεύτερη φάση τα πεπτίδια
είναι ικανά να αλληλοεπηρεάσουν το ένα με
το άλλο ,έτσι ώστε η έκκριση του ενός να
θεωρείται ως διεγέρτης για την έκκριση
κάποιου άλλου. Όπως θα δούμε στη συνέχεια στη κεφαλική φάση των γαστρικών
εκκρίσεων όπου δεν έχει φτάσει ακόμη η τροφή στο στομάχι η έναρξη παραγωγής του
γαστρικού οξέος γίνεται ως αποτέλεσμα διέγερσης του πνευμονογαστρικού.


Η έκκρισή γαστρικού οξέος ρυθμίζεται
με νευρικούς-νευροκρινικούς ,παρακρινικούς και ορμονικούς μηχανισμούς.
Παράγοντες που συμβάλουν στην έκκριση
γαστρικού οξέος είναι οι ακόλουθοι.
§ Το
Πνευμονογαστρικό .Οι απαγωγές ίνες του
πνευμονογαστρικού δεν νευρώνουν απ’ ευθείας τα εμβόλιμα ή τα νευροενδοκρινικά κύτταρα
συνάπτονται με μεταγαγγλιακούς νευρώνες του ΕΝΣ με μια ποικιλία νευροδιαβιβαστών
όπως Ach,GRP,VIP και PACAP κοντά
στην υποβλεννογόνια στοιβάδα. Οι μεταγαγγλιακοι νευρώνες έχουν την άμεσή
ικανότητα δράσης χρησιμοποιώντας Ach αν πρόκειται για
χολινεργικούς νευρώνες είτε έμμεσης
χρησιμοποιώντας κάποιο νευροενδοκρινές (Πεπτιδεργικοί νευρώνες)
§ Η
Ισταμίνη που παράγεται από τα ECL cells δρα στα εμβόλιμα κύτταρα προκαλώντας την
έκκριση του γαστρικού οξέος. Η Ισταμίνη θεωρείται ο κύριος διεγέρτης παραγωγής HCl .Θα αναλυθεί στη
συνέχεια.
§ Η
Γαστρίνη (ορμόνη)εισέρχεται στη κυκλοφορία
του αίματος με σκοπό να μεταφερθεί στα εμβόλιμα κύτταρα .
Ως
δευτερεύοντες παράγοντες μπορεί να θεωρηθούν:
§ Η
Ινσουλίνη με παρόμοιο τρόπο μεταφοράς με αυτό της Γαστρίνης φτάνει στα εμβόλιμα κύτταρα και προκαλώντας την έκκριση
HCl.
Ανεπαρκής ποσότητα γλυκόζης στον εγκέφαλο μπορεί να ενεργοποιήσει το μηχανισμό
§ Η καφεΐνη ως phosphodiesterase inhibitor αυξάνει τη συγκέντρωση
του cAMP στα εμβόλιμα κύτταρα αυξάνοντας με το τρόπο αυτό τη δράση
της αντλίας H+ - K+
ATPase .
§ Το στρες αυξάνει την έκκριση
του γαστρικού οξέος σε ορισμένα άτομα με μηχανισμό που δεν είναι απόλυτα
κατανοητός.
Σημαντικό ρόλο στην έκκρισή του γαστρικού οξέος διαδραματίζουν ορισμένα
γαστρεντερικά πολυπεπτίδια.
Τα γαστρεντερικά πολυπεπτίδια ( είτε προέρχονται από νευρικές απολήξεις
όπως η Ακετυλεχολίνη είτε από ενδοκρινή
κύτταρα , Ισταμίνη και Γαστρίνη
)συμβάλουν με 2 παράλληλους αλλά αλληλοσυνδεόμενους μηχανισμούς στην έκκριση του γαστρικού οξέος.
Σε πρώτη φάση και τα τρία πεπτίδια συνδέονται το κάθε ένα με ειδικό υποδοχέα
για στη βασεοπλευρική μεμβράνη του εμβόλιμου
κυττάρου (parietal cell) .
Οι απαγωγές νευρικές ίνες του
πνευμονογαστρικού συνάπτονται είτε με χολινεργικούς νευρώνες χρησιμοποιώντας ως νευροδιαβιβαστή την
Ακετυλεχολίνη (ΑCh) είτε με πεπτιδεργικούς peptidendric νευρώνες χρησιμοποιώντας ως νευροδιαβιβαστή
νευροπεπτιδia όπως (GRP, VIP και PACAP).
Το πνευμονογαστρικό είτε μέσο της ACh είτε με τη βοήθεια της Μπομπεσίνης (bombesin-gastrin-releasing peptide GRP) προκαλεί
την έκκριση της γαστρίνης από τα G κύτταρα
του πυλωρικού άνδρου.
Η οποία στη συνέχεια θα περάσει στη κυκλοφορία του αίματος (ορμόνη) θα
ενωθεί με τον υποδοχέα της στο εμβόλιμο κύτταρο
με σκοπό την έκκριση γαστρικού οξέος και πεψινογόνων
Πέρα από αυτό η έκκριση ήδη
Ακετυλεχολίνης και Γαστρίνης διεγείρουν την έκκριση της Ισταμίνης από τα
εντεροχρωμιόφιλα κύτταρα (ECL cells )συμβάλλοντας με έμμεσο τρόπο στην
έκκριση μεγαλύτερης ποσότητας HCl.
Όπως θα δούμε στην συνέχεια ο κύριος αναστολέας της έκκρισης του
γαστρικού οξέος είναι η Σωματοστατίνη SST.Το πνευμονογαστρικό
αναστέλλει τη δράση της Σωματοστατίνη για το χρονικό διάστημα που απαιτείται η
έκκριση του γαστρικού οξέος. Στη περίπτωση αυτή
η Ακετυλεχολινή ενώνεται με τους
Μ2-Μ4 υποδοχείς για αυτή που βρίσκονται
στην επιφάνεια του κυττάρου D με
σκοπό την παρεμπόδιση έκκρισης SST.
Όπως προαναφέρθηκε για καθένα γαστρεντερικό πεπτίδιο υπάρχει και ένας
ξεχωριστός υποδοχέας στη κυτταρική μεμβράνη του εμβόλιμου κυττάρου.
Πιο συγκεκριμένα :
§
Η
Ακετυλεχολίνη ενώνεται με τους Μ3 Μουσκαρινικούς υποδοχείς
§
Η Γαστρίνη
συνδέεται με τους cholecystokinin B υποδοχείς CCKB
Για
την Ισταμίνη υπεύθυνοι είναι οι H2 υποδοχείς
H Ακετυλεχολίνη
που εκκρίνεται από τις νευρικές απολήξεις του πνευμονογαστρικού συνδέεται με
τον μουσκαρινικό Μ3 υποδοχέα. Ας δούμε αναλυτικότερα
τι συμβαίνει Στον Μ3 υποδοχέα έρχεται να
ενωθεί μια πρωτεΐνη G GTP-binding protein (Gaq).
H Gaq πρωτεΐνη αποτελείται από 3 υπομονάδες
την aq , την β και την γ .Η αq unit ενεργοποιείται
από το GTP το
οποίο προσφέρει ένα φωσφορικό άλας σε αυτή phosphate και
μετατρέπεται σε GDP. Η αq εφόσον έχει ενεργοποιηθεί (switch on) αποκόπτεται από τις υπόλοιπες υπομονάδες
και συνδέεται με μια άλλη πρωτεΐνη στη κυτταρική μεμβράνη τη PLC phosphorolipase C.
Η σύνδεση αυτή ενεργοποιεί
την PLC η οποία στη συνέχεια θα δράσει σε μια νέα πρωτεΐνη
την PIP2. H σύνδεση της PLC με
την PIP2 θα έχει σαν αποτέλεσμα
τη διάσπαση της σε τριφωσφορική ινοσιτόλη inositol 1,4,5-triphosphate (IP3) και διακυλογλυκερόλη diacylglycerol (DAG).
Τόσο η ΙΡ3 όσο και η DAG αυξάνουν τη κυτταρική δραστηριότητα
και τη ενδοκυτταρική ποσότητα του Ca+2.
H IP3 θα δράσει στο ενδοπλασματικό δίκτυο όπου υπάρχουν αντλίες IP3 (ΙP3 dependent pumps). Η είσοδος IP3 στο ενδοπλασματικό δίκτυο έχει σαν αποτέλεσμα την έξοδο από
αυτό και προς το κυτταρόπλασμα ιόντων ασβεστίου.
Τα ιόντα αυτά στη συνέχεια θα δράσουν
σε μια πρωτεΐνη τη calmodulin ουσιαστικά μια πρωτεΐνη που η λειτουργία της ρυθμίζεται από τη παρουσία ασβέστιου. Η ένωση ιόντων ασβεστίου και της calmodulin δημιουργεί ένα calcium calmodulin complex που στη συνέχεια
θα δράσει σε μια συγκεκριμένη πρωτεϊνική κινάση CMK .Η οποία θα επιφέρει διάφορες
ενδοκυτταρικές αλλαγές και φυσικά θα συμβάλει και στην ενεργοποίηση της αντλίας
Η+ -Κ+.
Η DAG στο κυτταρόπλασμα θα ενεργοποιήσει μια πρωτεινική κινάση της PKC(πρωτεϊνική κινάση C) .Ένας από τους 3 ρόλους της PCK είναι
και η περαιτέρω ενεργοποίηση πρωτεϊνικών κινασών.
Η δράση
της ακετυλεχολίνης στο άνδρο του στομάχου
πέρα από τα G cells προσανατολίζεται και
στα D cells προκαλώντας αναστολή της έκκρισης
της Σωματοστατίνης .Η έκκριση Σωματοστατίνης θα μπλόκαρε την έκκριση Γαστρίνης .
Όταν η οξύτητα στον αυλό του στομάχου ξεπερνά μια
συγκεκριμένη τιμή αυτό έχει σαν αποτέλεσμα την ενεργοποίηση της Σωματοστατίνης προκαλώντας
αναστολή έκκρισης γαστρίνης και κατά συνέπεια περιορίζει την έκκριση γαστρικού οξέος(negative feedback mechanism).Αντίστοιχη ανασταλτική
δράση με αυτή της Σωματοστατίνης ασκούν
και προσταγλαδίνες. Ανασταλτική δράση στην έκκριση της Σωματοστατίνης STT ασκεί και η Ισταμίνη .Τα κύτταρα D του σώματος διαθέτουν
Η3 υποδοχείς ισταμίνης
.
Η Γαστρίνη (ορμόνη) εκκρίνεται
από τα G κύτταρα
και μέσω της κυκλοφορίας του αίματος καταλήγει στα εμβόλιμα κύτταρα όπου συνδέεται με τους CCKB gastrin-cholecystokinin B (CCKB)
receptor υποδοχείς. Ουσιαστικά 2 ξεχωριστοί υποδοχείς
έχουν αναγνωριστεί για την Γαστρίνη οι CCKA ή CCK1και οι CCKB ήCCK2 υποδοχείς . Οι CCKB μπορούν να δράσουν ταυτόχρονα και για τη Γαστρίνη αλλά η κύριά
δράση τους επικεντρώνεται στην CCK, ενώ οι CCKA υποδοχείς είναι αποκλειστικοί
υποδοχείς για τη CCK .
Όπως
και στη περίπτωση της Ach έτσι και οι υποδοχείς CCKB ενώνονται με G
GTP-binding protein (Gaq) και το ίδιο κυτταρικό μονοπάτι ακολουθείται (ενεργοποίηση PLC, που στη συνέχεια και μετά από συγκεκριμένα στάδια
θα ενεργοποίηση πρωτεϊνικες κινάσες είτε μέσω του IPP3 είτε του DAG και κατά συνέπεια την
έκκριση Ca2+.
Η έκκριση
της είναι αποτέλεσμα διάτασης του στομάχου , επίδραση του πνευμονογαστρικου. H παρουσία πεπτιδίων –αμινοξέων στο γεύμα
ασκούν ένα (positive feedback mechanism) στην
έκκριση της γαστρίνης και κατά επέκταση στην έκκριση γαστρικού οξέος.
Το Ασβέστιο αποτελεί διεγέρτη για την έκκριση γαστρίνης έτσι πολλοί ασθενείς με έλκος παρουσιάζουν
χρόνια υπερασβεσταιμία.
H γαστρίνη
με δυο διαφορετικούς τρόπους αυξάνει την έκκριση του γαστρικού οξέος είτε με
την διέγερση του εμβόλιμου κυττάρου, είτε με τη διέγερση των εντεροχρωμιόφιλων
κυττάρων προκαλώντας την έκκριση της ισταμίνης.
Η
SST και η interleukin-1β
αναστέλλουν την έκκρισης της γαστρίνης.
Η Ισταμίνη απελευθερώνεται από τα εντεροχρωμιόφιλα κύτταρα
enterochromaffin cells (ECL). Οι υποδοχείς της στη βασεοπλευρική μεμβράνη του εμβόλιμου κυττάρου
είναι οι Η2 υποδοχείς. Οι Η2 υποδοχείς ενώνονται στη συνέχεια
με μια διαφορετική GTP binding protein και πιο συγκεκριμένα
την Gas (Stimulatory) protein.H ένωση
αυτή θα έχει σαν αποτέλεσμα την τελική ενεργοποίηση πρωτεϊνικών κινασών (PKA) μέσω διαφορετικής οδού.
Η ενεργοποίηση του ενζύμου adenylyl cyclase (AC). στη συνέχεια θα οδηγήσει στην αύξηση της ενδοκυττάριας
ποσότητας cAMP. και ενεργοποίηση της PKA , η οποία θα προβεί
σε φωσφορυλίωση ειδικών πρωτεϊνών στο εμβόλιμο κύτταρο ,οι οποίες θα θέσουν σε λειτουργία
την αντλία ιόντων.
Διεγερτικά της
υποφυσιακής αδενυλικής κυκλάσης πεπτίδια -Pituitary adenylate cyclase-activating polypeptide- PACAL
Πρόκειται για
ένα νευροενδοκρινές που ανήκει στην ομάδα secretin–glucagon–VIP family. Το PACAP διαθέτει τρεις υποδοχείς . Τον
PACAP receptor type-1 (PAC-1), VPAC-1
(ο κλασικός VIP receptor),
και τον VPAC-2.
Ο PAC-1
παρουσιάζει μεγαλύτερη χημική συγγένεια για το PACAP
και μικρότερη για το VIP. Ενώ οι δυο άλλοι
υποδοχείς VPAC-1
και VPAC-2
παρουσιάζουν ισάξια ικανότητα-επιλογή
σύνδεσης τόσο με το PACAP όσο και το VIP.
Στο στομάχι έχουν ανακαλυφτεί PAC-1 receptors στα ECL κύτταρα , όπου η σύνδεση
του PACAP με τον PAC-1 receptor έχει
σαν αποτέλεσμα την αύξηση έκκριση της ισταμίνης .VPAC-1 receptor διαθέτουν τα D κύτταρα του σώματος του
στομάχου .Η σύνδεση του PACAP με τον VPAC-1 receptor έχει σαν αποτέλεσμα
την έκκριση σωματοστατίνης SST.
Τα D κύτταρα του πυλωρικού
άνδρου διαθέτουν επίσης VPAC-1 receptor με τη διαφορά ότι σε αυτόν συνδέεται η VIP με
αποτέλεσμα και πάλι την έναρξη έκκρισης σωματοστατίνης.
Το PACAP ουσιαστικά
διαθέτει την ικανότητα είτε να πυροδοτεί είτε να αναστέλλει την έκκριση
του γαστρικού οξέος.
ΑΛΕΞΑΝΔΡΟΣ ΜΟΥΜΤΖΗΣ